
Introdução

Um dos principais problemas dos seres vivos é a obtenção de energia para as suas atividades. De acordo com a teoria heterotrófica, os primeiros seres vivos seriam procariontes heterotróficos vivendo num meio aquático, donde retirariam nutrientes, formados na atmosfera e acumulados nos lagos e oceanos primitivos.
Devido á sua grande simplicidade, estes seres utilizariam processos igualmente rudimentares de retirar energia dessas moléculas de que se alimentavam. Esse mecanismo seria, quase com certeza, semelhante á fermentação realizada ainda por muitos organismos atuais.
Há mais de 2 mil milhões de anos, deverão ter surgido os primeiros organismos autotróficos, procariontes ainda mas capazes de produzir o seu próprio alimento através da fotossíntese. Este processo revolucionário, além de permitir a sobrevivência dos autotróficos, também serviu os heterotróficos, que passaram a alimentar-se deles.
A fotossíntese levou á acumulação de oxigênio na atmosfera terrestre, permitindo a alguma estirpes de procariontes tirarem partido do poder oxidante dessa molécula para retirar muito mais energia dos nutrientes, através da respiração.
Os organismos retiram energia das mais diversas moléculas orgânicas (açucares, aminoácidos, ácidos gordos, etc.), mas a glicose é a mais freqüente, tanto na fermentação como na respiração.

Fermentação
A fermentação é um conjunto de reações químicas controladas enzimaticamente, em que uma molécula orgânica (geralmente a glicose) é degradada em compostos mais simples, libertando energia. Este processo tem grande importância econômica, sendo utilizado no fabrico de bebidas alcoólicas e pão, entre outros alimentos.
Estudos realizados por Pasteur permitiram verificar que a fermentação alcoólica estava sempre associada ao crescimento de leveduras, mas que se estas fossem expostas a quantidades importantes de oxigênio produziriam (em vez de álcool e dióxido de carbono) água e dióxido de carbono. Destas observações, Pasteur concluiu que a fermentação é o mecanismo utilizado pelos seres vivos para produzir energia na ausência de oxigênio.
Já em 1897, o químico alemão Buchner demonstrou que a fermentação era apenas uma seqüência de reações químicas, podendo ocorrer fora de células vivas. Foi este estudo que revelou as enzimas (enzima = na levedura) e permitiu a compreensão do metabolismo celular em toda a sua globalidade.
Em 1930 os bioquímicos alemães Embden e Meyerhof descobriram a totalidade das etapas deste processo, pelo que essa seqüência também é conhecida por cadeia de Embden-Meyerhof.
Dependendo do tipo de microrganismo presente, a fermentação pode ser:
- Fermentação alcoólica - produz como produtos finais etanol e dióxido de carbono, produtos utilizados pelo Homem na produção de vinho, cerveja e outras bebidas alcoólicas e do pão (feita por fungos);
- Fermentação acética - produz como produto final o ácido acético, que causa o azedar do vinho ou dos sumos de fruta e sua conseqüente transformação em vinagre (feita por bactérias);
- Fermentação láctica - produz como produto final o ácido láctico, geralmente a partir da lactose do leite. O baixar do pH causado pela acumulação do ácido láctico causa a coagulação das proteínas do leite e a formação do coalho usado no fabrico de iogurtes e queijos (feita por bactérias).
 Pode-se considerar as reações da fermentação divididas em duas partes principais: a glicólise e a redução do ácido pirúvico.
Pode-se considerar as reações da fermentação divididas em duas partes principais: a glicólise e a redução do ácido pirúvico.

A glicólise é o conjunto de reações iniciais da degradação da glicose, semelhantes em todos os tipos de fermentação e na respiração aeróbia. Tem início com a ativação da glicose, que recebe dois grupos fosfato, fornecidos pelo ATP, que se transforma em ADP.
Por este processo de fosforilação a glicose transforma-se em frutose 1,6-difosfato (molécula com 6 carbonos e dois fosfatos) que será quebrada em duas moléculas de gliceraldeído 3-fosfato (molécula com 3 carbonos e um fosfato), pois é altamente instável.
A energia desta quebra permite a ligação de um outro grupo fosfato inorgânico a cada uma destas moléculas, que se tornam gliceraldeído 1,3-difosfato. Estes grupos fosfato, energéticos, são então transferidos para moléculas de ADP, transformando-as em ATP. O gliceraldeído transforma-se, por sua vez, em ácido pirúvico.

Sabe-se que a glicólise ocorre em praticamente todos os seres vivos, mesmo que complementada com outras reações, o que parece confirmar que deverá ter sido o primeiro fenômeno eficiente de produção de energia em células.
A segunda parte da fermentação consiste na redução do ácido pirúvico resultante da glicólise. Cada molécula de ácido pirúvico é reduzida pelo hidrogênio que é libertado pelo NADH2 produzido na glicólise, originando, conforme o tipo de organismo fermentativo, ácido láctico, ácido acético ou álcool etílico e dióxido de carbono.

RESPIRAÇÃO CELULAR

A fase aeróbica da respiração celular possui as seguintes etapas:
I - Formação da molécula de Acetilcoenzima A que irá entrar no Ciclo de Krebs.

II - Ciclo de Krebs ou do ácido cítrico (citrato)
Oxidação do Ácido Pirúvico
As moléculas de ácido pirúvico resultantes da degradação da glicose penetram no interior das mitocôndrias, onde ocorrerá a respiração propriamente dita. Cada ácido pirúvico reage com uma molécula da substância conhecida como coenzima A, originando três tipos de produtos: acetil-coenzima A, gás carbônico e hidrogênios.
O CO2 é liberado e os hidrogênios são capturados por uma molécula de NADH2 formadas nessa reação. Estas participarão, como veremos mais tarde, da cadeia respiratória.
Em seguida, cada molécula de acetil-CoA reage com uma molécula de ácido oxalacético, resultando em citrato (ácido cítrico) e coenzima A, conforme mostra a equação abaixo:
Em seguida, cada molécula de acetil-CoA reage com uma molécula de ácido oxalacético, resultando em citrato (ácido cítrico) e coenzima A, conforme mostra a equação abaixo:
1 acetil-CoA + 1 ácido oxalacético 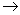 1 ácido cítrico + 1 CoA
1 ácido cítrico + 1 CoA
(2 carbonos) (4 carbonos) (6 carbonos)
(2 carbonos) (4 carbonos) (6 carbonos)
Analisando a participação da coenzima A na reação acima, vemos que ela reaparece intacta no final. Tudo se passa, portanto, como se a CoA tivesse contribuído para anexar um grupo acetil ao ácido oxalacético, sintetizando o ácido cítrico.
Cada ácido cítrico passará, em seguida, por uma via metabólica cíclica, denominada ciclo do ácido cítrico ou ciclo de Krebs, durante o qual se transforma sucessivamente em outros compostos.
Cada ácido cítrico passará, em seguida, por uma via metabólica cíclica, denominada ciclo do ácido cítrico ou ciclo de Krebs, durante o qual se transforma sucessivamente em outros compostos.
Analisando em conjunto as reações do ciclo de Krebs, percebemos que tudo se passa como se as porções correspondentes ao grupo acetil, anteriormente transferidas pela CoA, fossem expelidas de cada citrato, na forma de duas moléculas de CO2 e quatro hidrogênios. Um citrato, sem os átomos expelidos, transforma-se novamente em ácido oxalacético.
Os oito hidrogênios liberados no ciclo de Krebs reagem com duas substâncias aceptoras de hidrogênio, o NAD e o FAD, que os conduzirão até as cadeias respiratórias, onde fornecerão energia para a síntese de ATP. No próprio ciclo ocorre, para cada acetil que reage, a formação de uma molécula de ATP.
No esquema a seguir cada letra C representa um átomo de carbono.

Resumindo o Ciclo de Krebs

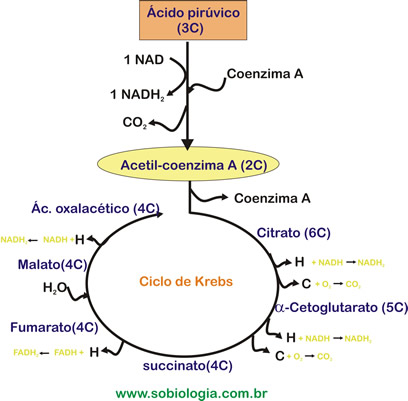
III - Cadeia respiratória - decorre na membrana interna da mitocôndria e consiste na transferência de 12 átomos de hidrogênio, libertados durante a oxidação da glicose, para o oxigênio. Esta transferência forma água e liberta energia. Em condições não celulares a libertação de energia seria explosiva mas este mecanismo gradual permite que esta seja utilizada. Cada conjunto completo de moléculas receptoras intermédias de hidrogênio (por vezes apenas o seu elétron, ficando o próton em solução) designa-se, então, cadeia respiratória. Além das moléculas de NAD e FAD, já referidas anteriormente, são fundamentais nesta cadeia os citocromos. De cada vez que um elétron é transferido há libertação de energia, mas apenas se forma ATP quando a energia é superior a 10000 calorias. Por vezes, a energia é suficiente para formar mais que uma molécula de ATP, mas apenas uma é sintetizada. O oxigênio, aceptador final de elétrons, fica carregado negativamente e combina-se com os prótons em solução, originando água.

A câmara de gás da morte

A Câmara de
gás é um dos suportes
utilizados pela justiça para execução de condenados a morte.
Foi criada nos EUA,
inspirado no uso de gases tóxicos durante a Primeira Guerra
Mundial.
O primeiro condenado à morte
a ser executado na câmara de gás foi um chinês de nome Gee Jon, no estado
norte-americano de Nevada, em 1924.
Na Alemanha nazista na II
Guerra Mundial essas câmaras eram muito solicitadas, em campos de
extermínio (em Varsóvia) do lado polonês na eliminação
sistemática de seus prisioneiros.
Nestas câmaras,
hermeticamente vedadas, um poderoso e mortal gás chamado Zyklon B era
injetado em quantidades no interior. O "Zyklon B" era o nome
comercial, mas na verdade, tratava-se do ácido cianídrico um gás
muito utilizado até hoje nas câmaras de gás norte americanas. O ácido
cianídrico usado para esse fim é uma pastilha forma cristais que uma vez
exposto ao ar entra em processo de sublimação e após algumas horas começa a
liberar o gás mortífero e altamente letal quando inalado. Para se ter uma
ideia, mesmo em pequenas doses, ao ser
respirado o gás cianídrico entra pela corrente sanguínea, até chegar
às células, onde bloqueia a ação das mitocôndrias, e desse modo as células
ficam sem produzir energia, ocorrendo a seguir a morte por asfixia. O gás também é usado em
grandes celeiros na eliminação do caruncho (pragas).
pt.wikipedia.org/wiki/C%C3%A2mara_de_g%C3%A1s
RESUMO ESQUEMÁTICO DA RESPIRAÇÃO CELULAR

Contabilidade energética da respiração aeróbica
Na glicólise há um rendimento direto de duas moléculas de ATP por moléculas de glicose degradada. Formam-se, também, duas moléculas de NADH2 que, na cadeia respiratória, fornecem energia para a síntese de de seis moléculas de ATP.
Durante o ciclo de Krebs, as duas moléculas de Acetil-CoA levam a produção direta de duas moléculas de ATP. Formam-se, também, também, seis moléculas de NADH2 e duas moléculas de FADH2 que, na cadeia respiratória, fornecem energia para a síntese de dezoito moléculas de ATP (para o NAD) equatro moléculas de ATP (para o FAD).
A contabilidade energética completa da respiração aeróbica é, portanto: 2 + 6 + 6 + 2 + 18 + 4 = 38 ATP. O resumo de todas as etapas resulta na seguinte equação geral:
1 C6H12O6 + 6 O2 + 38 ADP + 38 P 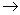 6 CO2 + 6 H2O + 38 ATP
6 CO2 + 6 H2O + 38 ATP
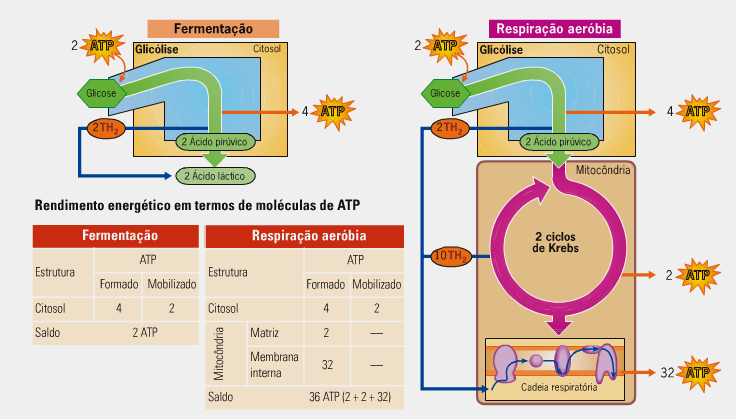

Observação 1) Em células musculares e neurônios tem-se observado um saldo de 36 ATP. Acredita-se que para os 2 NADH+H formados na glicólise entrarem na mitocôndria há um gasto de 2 ATP, Logo dos 38 ATP desconta-se 2ATP, restando 36 ATP.
Observação 2) Atualmente o saldo total é de 30 ou 32 ATP's, pois cada NADH+H gera 2,5 ATP e cada FADH2 gera 1,5 ATP, assim 10 NAD x 2,5 (25 ATP) + 2 FADH2 x 1,5 (3 ATP) + 2 ATP (glicólise) + 2 ATP (ciclo de Krebs) = 32 ATP.
Eficiência da Respiração
A glicose contem em termos de energia química, cerca de 685 000 calorias por mol. Já os ATP que ela permite reconstituir significam algo ao redor de 300 000 calorias. O rendimento da respiração aeróbia é portanto, de 44% aproximadamente, isso que dizer que uns 56% da energia existente na glicose se perdem durante as diversas etapas do processo. Ainda assim a respiração é considerada um processo bastante eficiente em termos energéticos. Motores a combustão, na melhor das hipóteses não atingem um rendimento de 25%.
http://www.tudomaisumpouco.com/aula6respira%C3%A7%C3%A3o.html
FONTES NUTRICIONAIS PARA RESPIRAÇÃO CELULAR
A nossa alimentação nos fornece carboidratos (ricos em glicose), proteínas (ricas em aminoácidos) e gorduras (ricas em ácidos graxos).
Acetil-CoA é formado a partir de Piruvato e da betaoxidação de Ácidos Graxos.

Respiração anaeróbia
Um restrito grupo de bactérias utilizam
compostos inorgânicos como aceptor final de elétrons
• O aceptor final de elétrons é uma
substância inorgânica diferente de oxigênio. Geralmente substâncias oxidadas:
NITRATOS, SULFATOS ECARBONATOS.
Bactérias
tais como:
• Pseudomonas e Bacillus podem utilizar o íon nitrato (NO3-)
como aceptor final de elétrons. O íon nitrato é reduzido a íon nitrito (NO2-), oxido nitroso (N2O) ou
gás nitrogênio (N2).
• Desulfovibrio e Desulfotomaculum utilizam sulfatos (SO42-) como
aceptor final de elétrons para formar sulfeto de hidrogênio (H2S).
• Bactérias metanogênicas utilizam carbonato (CO32-) para formar metano (CH4).
Rendimento: mais baixo que a respiração aeróbica,
somente uma parte do TCA funciona sob condições anaeróbias e
nem todos os transportadores participam da cadeia.
http://www.scribd.com/doc/17919137/Resp-Anaerobic-A-Fermentacao
Embora a Respiração Aeróbia seja mais usada entre
os seres vivos, existem alguns que não utilizam oxigênio livre durante este processo, o que recebe o nome de Respiração
Anaeróbia. Mas o que é Respiração anaeróbia, você sabe???
Como já foi dito a principal mas não única diferença entre a respiração aeróbia e a anaeróbia é que a primeira necessita de oxigênio do ambiente e a segunda não. O
processo anaeróbio pode ser praticado por algumas espécies de bactérias que liberam a energia química dos alimentos.
Ele necessita de substâncias inorgânicas que tenham oxigênio em sua molécula. Preste atenção, o fato de conter oxigênio na substância inorgânica não torna o
processo aeróbio, já que este necessita de oxigênio livre.
Essas substâncias podem ser nitratos, sulfatos ou carbonatos.
Confira um exemplo de reação de respiração aeróbia onde é utilizado o nitrato:

O nitrato é usado no processo de respiração anaeróbio de bactérias conhecidas como desnitrificantes do solo. Um exemplo é a espécie Pseudomonas denitrificans que participa do ciclo no nitrogênio.
A FOTOSSÍNTESE
FAVOR ACESSAR ESSAS ANIMAÇÕES IICIALMENTE:
http://www2.ib.unicamp.br/lte/bdc_uploads/materiais/versaoOnline/versaoOnline1140_pt/S_2_2_14_Fotossintese/Fotossintese.swf
http://www.profdorival.com.br/resources/Fotossintese.swf
http://www2.ib.unicamp.br/lte/bdc_uploads/materiais/versaoOnline/versaoOnline1140_pt/S_2_2_14_Fotossintese/Fotossintese.swf
http://www.profdorival.com.br/resources/Fotossintese.swf
A fotossíntese fornece alimento a todas as formas de vida, pois os organismos heterotróficos se alimentam direta ou indiretamente das moléculas orgânicas produzidas pelos autotróficos.
Outro importante contribuinte da fotossíntese é a produção de oxigênio, utilizado na respiração pela maioria dos organismos atuais. Praticamente todo o oxigênio da atmosfera terrestre tem origem fotossintética e pensa-se que é totalmente renovado, pelo mesmo processo, a cada 2000 anos.
A descoberta deste fenômeno fundamental para a vida na Terra é, apesar de tudo, bastante recente, tendo sido mencionado pela primeira vez em 1772 pelo inglês Priestley. Este bioquímico apercebeu-se que a introdução de uma planta num ambiente irrespirável melhorava rapidamente a qualidade do ar.
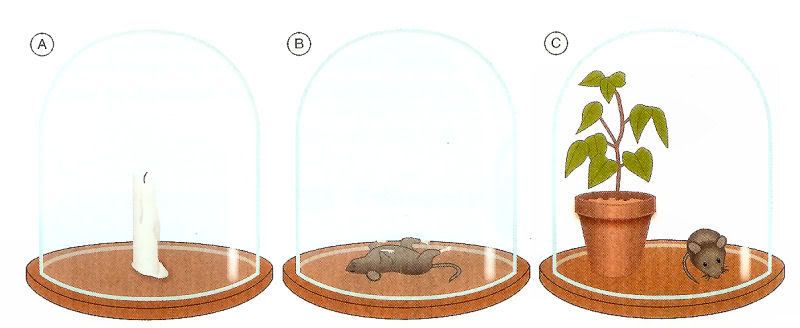
Experiência de Priestley que demonstrou que a chama de uma vela se apaga dentro de uma campânula (A); o rato morre dentro de uma campânula (B) e o rato sobrevive se estiver acompanhado de uma planta dentro da campânula (C).
In: Carrajola, C. (2007b).
Em 1779 o holandês Ingen-Housz notou que para que as plantas “recuperassem” o ar necessitavam de luz e que essa “recuperação” se devia a um enriquecimento do ar em oxigênio. Iniciou-se aqui a idéia que as plantas decompunham o dióxido de carbono, libertando oxigênio, embora não fosse claro o destino do carbono excedente.
O mesmo Ingen-Housz propôs em 1796 que as plantas o utilizavam para fabricar as suas próprias moléculas orgânicas, sendo o oxigênio um subproduto dessas reações. A partir deste momento, o mecanismo ficou batizado fotossíntese (síntese em presença de luz de compostos orgânicos).

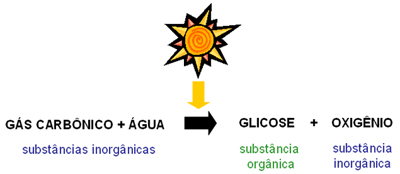
As complexas reações da fotossíntese ocorrem nos cloroplastos, organitos semi-autónomos presentes nos seres autotróficos, e podem ser resumidas da seguinte forma:

Esta forma de resumir a fotossíntese, embora correta, não revela a complexidade das reações intermédias e dá a idéia (errada) de que o dióxido de carbono reage com a água.
Por volta de 1930, o investigador Van Niel propôs a hipótese que o oxigênio libertado na fotossíntese proviesse da água e não do dióxido de carbono, como antes se pensava. Dez anos mais tarde experiências com isótopos pesados de oxigênio comprovaram esse fato.

Outro tipo de experiências revelou que algumas das reações da fotossíntese são fotoquímicas (realizam-se em presença de luz), enquanto outras são termoquímicas (realizam-se sem intervenção direta da luz). Assim, é regra dividir o processo em fase luminosa, que ocorre a nível dos grana do cloroplasto, e fase escura, cujas reações decorrem no estroma.


A luz é constituída por “partículas luminosas”, altamente energéticas, designadas fótons. A cor da luz é determinada pela energia dos fótons que a compõem (zona azul do espectro mais energia e zona vermelha do espectro menos energia).

No caso das reações da fotossíntese, as principais moléculas envolvidas são as clorofilas. Quando moléculas de clorofila são atingidas por luz de cor azul e vermelha (fótons com determinada energia, portanto), alguns dos seus elétrons passam a orbitais mais elevadas e a molécula fica excitada.
No entanto, a clorofila excitada é muito instável e ao fim de certo tempo os elétrons regressam ás suas órbitas de origem - estado fundamental -, libertando a energia que absorveram do fóton, sob a forma de luz. Este fenômeno é conhecido pela fluorescência da clorofila. As clorofilas refletem a luz verde, sendo esse o motivo porque as plantas são verdes.

Na maioria das células vegetais existem dois tipos de clorofila, a e b, sendo a clorofila b mais oxidada.

As moléculas de clorofila, receptores de elétrons, pigmentos acessórios e enzimas participantes na fotossíntese estão organizadas nas membranas dos cloroplastos em unidades designadas fotossistemas.
Cada fotossistema contém entre 250 a 400 moléculas de pigmentos e consiste em dois componentes intimamente associados: um centro de reação (formado por um complexo proteína-pigmento) e um complexo antena.
Todas as moléculas de pigmentos do fotossistema são capazes de absorver fóton, mas apenas um par de moléculas de clorofila em cada fotossistema utiliza essa energia nas reações fotoquímicas. Este par, localizado ao centro do fotossistema forma o centro de reação, enquanto as restantes moléculas se designam pigmentos antena. Estes podem ser, além de clorofilas, carotenoides e ficobilinas (ficocianina azul e ficoeritrina vermelho).
Dentro dos fotossistemas, as moléculas de pigmentos estão ligadas a proteínas específicas e situadas em locais que permitem uma eficiente captação da energia luminosa. A energia absorvida por cada molécula é transferida á seguinte, até alcançar o centro de reação. Quando ambas as clorofilas do centro de reação absorvem energia, um dos seus elétrons é excitado e transferido para a primeira molécula receptora, iniciando-se o fluxo de elétrons necessário ás reações fotoquímicas.
Existem dois tipos de fotossistemas:
Fotossistema I - também designado PS I, contém no seu centro de reação uma forma de clorofila a designada P700, pois absorve luz de comprimento de onda de 700 nm. Localiza-se preferencialmente nas membranas intergrana, em contacto direto com o estroma do cloroplasto;

Fotossistema II - também designado PS II, contém no seu centro de reação uma forma de clorofila a designada P680 (clorofila b), pois absorve luz de comprimento de onda de 680 nm. Localiza-se nos tilacóides e procede a fotólise da água.

De modo geral, os fotossistemas funcionam simultaneamente, mas o fotossistema I pode funcionar independentemente.

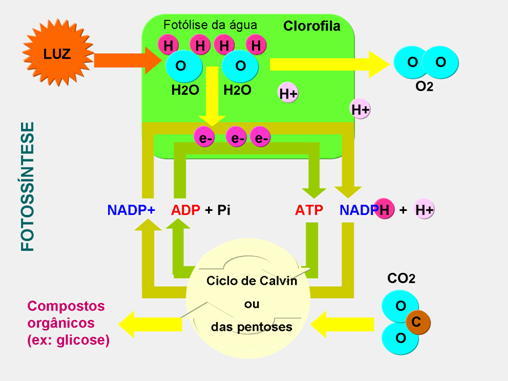
No interior da célula, a energia libertada pelo regresso do elétrons á sua orbital original não é “perdida” sob a forma de luz mas sim captada por um conjunto de moléculas, sendo depois utilizada na síntese de moléculas de ATP e NADPH2, utilizadas nas reações da fase escura.
A síntese destas moléculas implica dois tipos de reações:
Fotofosforilação acíclica - nesta reação já intervêm os dois tipos de clorofila a e, logo, ambos os fotossistemas. A molécula de clorofila P680 é excitada ao ser atingida por um fóton. Os seus elétrons libertam-se e são captados por um receptor de elétrons, a plastoquinona. Dessa molécula, os elétrons passam por outra cadeia transportadora de elétrons, perdendo energia, que é utilizada na síntese de ATP a partir de ADP e fosfato inorgânico. A última molécula dessa cadeia é uma clorofila P700 oxidada. Ao receber o elétron ficará, portanto, reduzida. No entanto, ao receber o estímulo de novo fóton, volta a perder o seu elétron excitado, que é passado á ferredoxina e dela para o NADP, que fica reduzido (NADP2).

O oxigênio produzido pela fotólise da água é eliminado para a atmosfera e os elétrons vão substituir os elétrons perdidos pela clorofila P680 durante a fotofosforilação acíclica, permitindo que regresse á sua forma reduzida. Os prótons H+ são captados pelo NADP2-, originando NADPH2.

Na fase escura da fotossíntese ocorrem uma série de reações com absorção e redução de dióxido de carbono, inversas da glicólise, com formação de compostos orgânicos (açúcares, aminoácidos, ácidos gordos, glicerol, etc.). No decorrer desta fase há gasto de NADPH2 e ATP, formadas na fase luminosa, as quais se transformam em NADP e ADP e voltam ás reações da fase luminosa.
Foram as experiências de Calvin, Bassham e Benson, entre 1954 e 1960, que permitiram determinar as diferentes etapas desta fase da fotossíntese. Por esse motivo, a série de reações que permitem a síntese de glicose a partir de dióxido de carbono, ATP e NADPH2 é conhecida por ciclo de Calvin-Benson ou ciclo das pentoses.
O produto primário do ciclo de Calvin é o gliceraldeído 3-fosfato, a molécula transportada do cloroplasto para o citoplasma da célula. Esta é exatamente a mesma molécula produzida pela quebra da frutose 1,6-difosfato na glicólise.
A enzima ribulose 1,5-difosfato carboxilase, vulgarmente designada Rubisco, a enzima catalisadora da reação inicial do ciclo de Calvin (fixação do dióxido de carbono na ribulose) é muito abundante nos cloroplastos, correspondendo a mais de 15% do seu conteúdo protéico total. É, por este motivo, considerada por muitos bioquímicos a proteína mais abundante do mundo.

Assim, os fenômenos da fotossíntese podem ser resumidos, considerando apenas os produtos iniciais e finais, da seguinte forma:
RESUMO GERAL DA FOTOSSÍNTESE

Embora a glicose seja a molécula representada nas equações reduzidas da fotossíntese, a quantidade de glicose livre produzida nas células fotossintéticas é muito baixa. A maioria do carbono fixado é convertido preferencialmente em sacarose, o glicídio de transporte, ou em amido, o glicídio de reserva, das plantas.
O gliceraldeído 3-fosfato que é transportado para o citoplasma da célula é utilizado para formar glicose 1-fosfato, precursor imediato da sacarose. Pelo contrário, o gliceraldeído 3-fosfato que permanece nos cloroplastos, passa a amido, armazenado sob a forma de grânulos no estroma. Durante a noite, a glicose do amido é exportada para o citoplasma.

A glicose é a base para vários outros carboidratos energéticos e estruturais.
FOTOSSÍNTESE BACTERIANA
- Cianobactérias: são fotolitoautotróficas e aparentemente foram as pioneiras no uso da água como fonte de elétrons. Incluiriam as proclorófitas (gêneros Prochloron, Prochlorothrix eProchlorococcus), apesar de se distinguirem destas por apresentar apenas clorofila a, além de ficobilinas azul e vermelha. Esses pigmentos são responsáveis pelas diversas colorações, muitas vezes brilhantes, que essas bactérias apresentam.
CO2 + H2O --> CH2O + O2
PRODUZ GLICIDIO E GÁS OXIGÊNIO
- Sulfobactérias: realizam um tipo de fotossíntese em que a substância doadora de hidrogênio não é a água, mas compostos de enxofre, principalmente o gás sulfídrico (H2S). Por isso essas bactérias produzem enxofre elementar (S) como subproduto da fotossíntese, e não gás oxigênio, como na fotossíntese que utiliza H2O.
CO2 + 2 H2S --> CH2O + 2S +
H2O
Observações:
Não possuem clorofilas vegetais a e/ou b.
Possuem clorofila do tipo Bacterioclorofila.
A Bacterioclorofila está dispersa no citoplasma pois não ocorre a presença de cloroplastos.
Não utilizam o H2O como fonte de O2 e sim o H2S.
Não liberam O2 e sim 2S.
Possuem clorofila do tipo Bacterioclorofila.
A Bacterioclorofila está dispersa no citoplasma pois não ocorre a presença de cloroplastos.
Não utilizam o H2O como fonte de O2 e sim o H2S.
Não liberam O2 e sim 2S.
QUIMIOSSÍNTESE
FOTOSSÍNTESE X QUIMIOSSÍNTESE
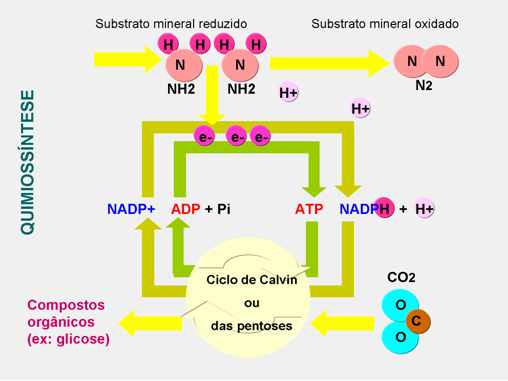
A quimiossíntese é a produção de matéria orgânica através da oxidação de substâncias minerais, sem recorrer à luz solar.
A quimiossíntese divide-se em duas etapas:
- A formação do NADPH e de ATP, usando a energia fornecida por determinadas reações químicas de oxirredução que ocorrem no meio;
- A segunda fase é igual à fase escura da fotossíntese: redução de dióxido de carbono, o que conduz à síntese de substâncias orgânicas.
A quimiossíntese é realizada principalmente por bactérias, entre as quais as ferrobactérias, as sulfobactérias e as nitrobactérias. Cada uma dessas bactérias utiliza a energia de um mineral que oxida, compostos respectivamente de ferro, enxofre e nitrogênio.
Comparada à fotossíntese, a taxa da quimiossíntese apresenta uma fração minúscula de produção, mas é muito importante para o ciclo do nitrogênio, onde este elemento é fixado no solo ou nas plantas.

